bioloske oksidacije i celijsko disanje
Bioloske oksidacije i celijsko disanje
Bioloske oksidacije i celijsko disanje su kljucni procesi u kojima celija razlaze organske molekule i oslobadja energiju za osnovne funkcije.
Celijsko disanje je proces biohemijske oksidacije, odnosno razgradnje organskih molekula, kao sto je glukoza, pri cemu se oslobadja energija potrebna za zivotne funkcije celije. To je niz reakcija koje se odvijaju unutar celije, tokom kojih se slozeniji molekuli razlazu na jednostavnije, oslobadjajuci energiju pohranjenu u hemijskim vezama.
Bioloske oksidacije i celijsko disanje
Kicmenjaci i ostale metazoe , kao i veliki broj mikroorganizama , spadaju u striktno aerobne organizme,tj. u organizme za ciji zivot neophodno prisustvo kiseonika .
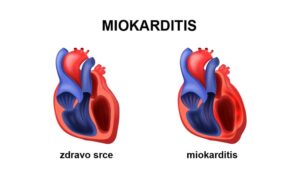
Metabolicke odlike srcanog misica 26
Metabolicke odlike srcanog misica Metabolicke odlike srcanog misica .Srcani misic (miokard) ima specificne metabolicke odlike...

Bioelektricne pojave u srcu 26
Bioelektricne pojave u srcu EKG elektrokardiogram Bioelektricne pojave u srcu omogucavaju stvaranje i sirenje elektricnih...

Fizioloske osobine srcanog misica 26
Fizioloske osobine srcanog misica Fizioloske osobine srcanog misica (miokarda)omogucavaju neprekidan rad srca kroz automatsko stvaranje...
U fiziologiji disanja videli smo da covek , kao i svi ostali kicmenjaci , dobija kiseonik iz atmosferskog vazduha koji procesom udisanja dospeo u plucne alveole .
Ako ste propustili :
Posredstvom krvi , vezan za hemoglobin eritrocita u obliku oksihemoglobina , kiseonik se prenosi do tkivnih kapilara i iz njih dospeva u celije , gde ucestvuje u oksidacionim procesima .
O sustini bioloskih oksidacija postala su razlicita shvatanja u nauci. Danas je nesumnjivo da se u toku procesa oksidacija u celijama , tj. bioloskih oksidacija , kiseonik jedini sa vodonikom iz organskih jedinjenja , pri cemu postaje voda i oslobadja se energija:
2 H +1/2 O2 —–> H2O + E
Bioloske oksidacije i celijsko disanje
Bioloske oksidacije i celijsko disanje . potrebno je imati na umu da atmosferski kiseonik , kao i vodonik organskih jedinjenja , nisu reaktivni i da je za pomenutu reakciju sinteze vode neophodno da vodonik bude oduzet od organskih jedinjenja i da vodnik i kiseonik budu aktivirani .
Bioloske oksidacije i celijsko disanje . Znaci gornja jednacina u stvarnosti izgleda nesto drugacije :
2 H+ + O— ——–> H2O + E
Na kraju treba istaci da se samoo jedan deo energije oksidacionih procesa oslobadja u obliku toplote , dok se znatan deo akumulira i konzervise , a zatim koristi za razne bioloske procese i zivotne manifestacije .
Aktiviranje kiseonika u celijskom disanju
Aktiviranje kiseonika u celijskom disanju se vrsi pod uticajem Varburgovog fermenta disanja ili cim-oksidaze .Osobine Varburgovog fermenta opisali smo ranije .
Ovde ponavljamo njegove najvaznije osobine : da je autooksidabilan , tj. u prisustvu molekulskog kiseonika e oksidise , pri cemu gvozdje njegovog koencima prelazi iz fero-oblika , u feri-oblik , dok se kiseonik prmanjem elektrona iz ovog gvozdja pa od molekulskog u aktivni.
Preporuka knjige :

Istorijski Atlas – kupite knjigu
Bioloske oksidacije i celijsko disanje . Ovaj proces se vrsi bez ikakvih drugih posrednika . Varburgov ferment je osetljiv prema dejstvu CO i HCN , koji ga inaktivisu .
Aktiviranje kiseonika u celijskom disanju . Redukcija Varburgovog fermenta vrsi se pod uticajem narocite grupe jedinjenja – prenosilaca elektrona , koji se nazivaju citohromi.
Vazno je podvuci da su citohromi feroprotoporfirini , koji nisu autooksitabilni i koji igraju ulogu u prenosenju elektrona u lancu oksidaciono-redukcionog procesa .
U bioloskim okidacijama u celijama coveka najcesce ucestvuju tri citohroma : a, b i c , koji funkcionisu odredjenim redom , prema prilozenoj semi :
4W F – Fe++ O2 ——–> 4 W F – Fe+++ +2O—
4 W F -Fe+++ + cyt a – Fe++ ———-> 4WF- Fe++ + 4 cyt a – Fe+++
4 cyt a Fe+++ + 4 cyt -c – Fe++ ———> 4 cyt a – Fe++ +4 cyt c -Fe+++
4 cyt -c -Fe+++ + 4 cyt -b -Fe++ (ili drugi prenosioci e–) ——-> 4 cyt -c–
-Fe++ + 4cyt -b-Fe+++ ( odnosno oksidisani drugi prenosioci elektrona (e– )
Aktiviranje kiseonika u celijskom disanju. Iz seme se vidi da Varburgov ferment , aktivisuci kiseonik predavanjem elektrona , postaje oksidan ( W F +Fe+++ ).
Aktiviranje kiseonika u celijskom disanju. Citohrom a redukuje Varburgov ferment predajuci mu elektron , pri cemu njegovo gvozdje prelazi u feri-oblik .
Aktiviranje kiseonika u celijskom disanju. Citrohrom c na isti nacin redukuje citrohrom a , dok njega redukuje bilo citrohrom b ili drugi prenosioci elektrona , o kojima se mogu naci podaci u biohemiji .
Aktiviranje kiseonika u celijskom disanju. Na kraju , fermenti dehidrogenaze jedinjenja ( glukoze, masnih kiselina , amino kiselina i dr. ), koje od organskih jedinjenja oduzimaju dehidrogenaze .
Aktiviranje kiseonika u celijskom disanju. Iz izlozenog se vidi da citohromi igraju ulogu prenosilaca elektrona sa vodonika oduzetog od supstrata ( pomenutih organskih jedinjenja ) do Vrburgovog fermenta .
Na kraju , redukuju Varburgov ferment , cime ga osposobljavaju za reagovanje sa novim molekulima kiseonika .
Aktiviranje kiseonika u celijskom disanju. Citohromi imaju jos jednu ne manje vaznu ulogu : pri prenosenju elektrona sa jednog citohroma na drugi dolazi do konzerviranja i akumuliranja znatnog dela energije u obliku fosfatnih jedinjenja bogatih energijom .

Aktiviranje vodonika u biohemijskim procesima
Aktiviranje vodonika u biohemijskim procesima . proces aktiviranja vodonika odigrava se pod uticajem femenata dehidrogenaza o kojima je u poglavlju o fermentima dosta bilo reci .
Ovde samo podsecamo da se prema strukturi koencima i nacinu funkcionisanja dehidrogenaze dele u dve grupe : a)nikotin-amid-dehidrogenaze i b) flavin-dehidrogenaze .
Prema funkciji se flavin-dehidroooze dele u dve grupe : prave flavin-dehidrogenaze , koje oksidisu nnnnganska jedinjenja ( npr. amino kiseline , vise masne kiseline ili cilibarnu kiselinu ) , i flavin-encimi – prenosioci elektrona , koji primaju aktivisane vodonike i elektrone od redukovanih nikotin-amid-dehidrogenaza i prenose ih na citohrome i druge prenosioce elektrona .
Ostali prenosioci elektrona
Prenosioci elektrona . Pored flavin-encima -prenoslica elektrona i citohroma , pominje se jos citav niz jedinjenja prenosilaca elektrona koji ucestvuju u oksidacionom lancu u celijama .
Prenosioci elektrona . Kao najvaznije od njih pominjemo : ubihinon (koencim Q) i vitamine E i K . Blizi podai o njihovim osobinama nalaze se u biohemiji .
Oksidacioni lanac u celijama
Oksidacioni lanac u celijama. O procesu prenosenja aktivisanog vodonika i elektrona sa organskih jedinjenja do aktivisanog kiseonika koji se zavrsava sintezom vode postoji jos dosta razlicitih shvatanja .
Oksidacioni lanac u celijama. Stavise , skoro je sigurno da ovaj proces nije istovetan u svim celijama ni pri oksidaciji svih jedinjenja , nego da su broj i vrsta prenosilaca elektrona narocito citohroma i posrednika – prenosilaca elektrona -razliciti , zavisno od toga koji je organizam , ili cak organ , po sredi i koje se jedinjenje oksidise.
Oksidacioni lanac u celijama. U biohemiji moze se naci vise podataka o ovom procesu i razlikama toka oksidacionog lanca , zavisno od organizma , organa ili jedinjenja .
Oksidacioni lanac u celijama. Ovde cemo se zadovoljiti time sto cemo daaato izmenjenu semu po Slateru , mada relativno prosta , pregledno prikazuje najvaznije etape oksidacionog lanca u organizmu kicmenjaka .
![\[ \Huge \renewcommand{\arraystretch}{1.5} \begin{array}{c} \boxed{\text{[A] NADH / FADH}_2} \\ \downarrow \\ \boxed{\text{[B] Flavoprotein (FMN/FAD)}} \\ \downarrow \\ \boxed{\text{[C] Koenzim Q (Ubihinon)}} \\ \downarrow \\ \boxed{\text{[D] Citohromi b, c_1, c}} \\ \downarrow \\ \boxed{\text{[E] Citohromi a, a_3} \rightarrow \text{ O}_2 \rightarrow \text{H}_2\text{O}} \\ \\ \text{★ P-sajt 1} \quad \text{★ P-sajt 2} \quad \text{★ P-sajt 3} \end{array} \]](https://prirodnolecenje.in.rs/wp-content/ql-cache/quicklatex.com-10df4f7ac485b5b68bdfff4c8dad0932_l3.png "Rendered by QuickLaTeX.com")
| Redni broj | Između kojih komponenti | ATP se stvara |
|---|---|---|
| P-sajt 1 | Između NADH (A) i flavoproteina (B) | ✅ |
| P-sajt 2 | Između koenzima Q (C) i citohroma b/c₁ (D) | ✅ |
| P-sajt 3 | Između citohroma c i citohroma a₃ (E) | ✅ |
Svaki od tih sajtova pokrece pumpanje protona (H⁺) kroz membranu → pravi se gradijent → koji ATP sintaza koristi za proizvodnju ATP-a.
Oksidacioni lanac u celijama. Iz seme se vidi da pri prenosenju vodonika i elektrona pod uticajem nikotin-amid-dehidrogenaza kao obavezni prenosilac ucestvuju flavin-enzimi prenosioci elektrona (diaforaza ili citohrom-c-reduktaza) koji elektrone predaju sistemu citohroma , dok flavin-dihidrogenaze nemaju nikakvih posrednika i elektrone direktno predaju citohromima.
Oksidacioni lanac u celijama. Bioloski znacaj ovako slozenog toka procesa prenosenja elektrona je u tome sto je velika kolicina energije koja se oslobadja pri sjedinjavanju aktivisanog vodonika sa aktivisanim kiseonikom (68.000 cal./mol) ne oslobadja se samo u obliku toplote , niti odjednom , nego u etapama , tako da nepostoji opasnost ostecenja zive materije naglim oslobadjanjem ovako velike kolicine toplote.
Oksidacioni lanac u celijama. Pored toga , znatan deo energije , do 50% ne oslobadja se u obliku toplote , nego se akumulira i konzervira u obliku adenozin-trifosfata ( ATF).
Fosfatna jedinjenja bogata energijom
Fosfatna jedinjenja bogata energijom . Proucavanje raznih jedinjenja fosforne kiseline , a narocito organskih , pokazalo se da se sva fosfatna jedinjenja mogu podeliti u dve grupe .
Fosfatna jedinjenja bogata energijom . U prvu grupu , ciji su predstavnici glicerih-aldehit-fosfat , glicerin-fosfat i adenilna kiselina , dolaze fosfatna jedinjenja male energije , za ciju sintezu nije potrebna velika kolicina energije , odnosno pri cijem se razlaganju ( odvajanju H3PO4) oslobadja mala kolicina energije ( ispod 3.000 cal./mol).
Fosfatna jedinjenja bogata energijom . U drugu grupu dolaze fosfatna jedinjenja bogata energijom , kao sto su adenozin-trifosfat , adenozin-difosfat , kreatin-fosfat i niz drugih , kod kojih se pri odvajanju jednog molekula fosforne kiseline oslobadja velika kolicina energije (8-11.000 cal./mol).
Fosfatna jedinjenja bogata energijom .Sinteza ovih jedinjenja je ondroenergijski proces tj. zahteva energiju spolja – iz nekog egzoenergijskog procesa koji se istovremeno odigrava .
Fosfatna jedinjenja bogata energijom . Detalji o biohimijskim reakcijama u organizmu pri kojima postaju fosfatna jedinjenja bogata energijom i potpuna lista fosfatnih jedinjenja bogatih energijom mogu se naci u biohemiji .
Ovde cemo se zadovoljiti time sto naglasavamo da je najvazniji proces koji daje energiju za vezivanje neorganske fosforne kiseline za adenozin-difosfat i sintezu adenozin-trifosfata vec pomenuti oksidacioni lanac .
Pri oksidacijama organskih jedinjenja pod uticajem nikotin-amid-enzima i prenosenju elektrona i vodonika do aktivisanog kiseonika energija se akumulira i sintetise ATF u tri etape :
flavin-cyt c , cyt c ->cyt b i cyt b -> Verburgov ferment. Na ovaj nacin se pri sintezi svakog molekula vode sintetisu tri molekula ATF.
Fosfatna jedinjenja bogata energijom .Adenozin-trifosfat ima vise uloga . predajuci molekul fosforne kiseline bogate energijom cija se veza obelezava ~ , omogucuje sintezu adenozin-difosfata od adenilne kiseline i kreatin-fosfata vezivanjem fosforne kiseline za kreatin..
Fosfatna jedinjenja bogata energijom . Adenozin-trifosfat daje energiju koja se neposredno koristi za niz vaznih fizioloskih procesa :
Regulisanje propustljivosti celicnih membrana , sintezu velikog broja sastojaka organizma (nukleinskih kiselina , belancevina , masti i dr.) , za misicnu kontrakciju , nervnu razdrazljivost ,procese rastenja itd.
Fosfatna jedinjenja bogata energijom . Kreatin-fosfat igra ulogu rezerve energije u celijama , a nnnnto u misicima .
On nije u stanju da svoju energiju neposredno stavlja celijama na koriscenje za fizioloske procese , nego se taj proces odigrava posredno : kreatin-fosfat predaje svoju fosfornu kiselinu bogatu energijom adenozin-difosfatu , pri cemu se najpre sintetise ATF , a ovo jedinjenje ucestvuje kao neposredni izvor energije .
Mehanizam dejstva fizioloski aktivnih supstancija – teorija o hemijskim receptorima celija
„Corpora non agunt nisi fixata“ – P.Ehrlich
Fosfatna jedinjenja bogata energijom . Jedna od najznacajnih odlika celija je da su osetljive prema dejstvu ogromnog broja hemijskih supstancija , koje po poreklu mogu biti :
- a) proizvodi samog organizma – proizvodi prometa materija , hemijski prenosioci (medijatori) nervnih impulsa , tkivni ili pravi hormoni
- b) bioloski proizvodi drugih organizma – biljnog , zivotinjskog i bakterijskog porekla
- c) veliki broj neorganskih , a narocito organskih sintetickih jedinjenja ( lekovi , otrovi ).
Fosfatna jedinjenja bogata energijom . Ove aktivne supstancije dejstvuju na celije na veoma razlicite nacine :
Uticu na propustljivost celijske opne , povecavaju ili smanjuju razdrazljivost , uticu na propustljivost u nervnom sistemu ,ubrzavaju , usporavaju ili cak potpuno koce metabolicke procese , dejstvuju na kvalitet i tok misicne kontrakcije , pojacavaju ili smanjuju sekreciju itd.
Fosfatna jedinjenja bogata energijom . Neke od tih supstancija mogu dovesti do trajnih poremecaja celija i ukoliko se radi o zivotno vaznim organima (srce , centralni nervni sistem ) , mogu dovesti i do smrti .
Fosfatna jedinjenja bogata energijom . Razumljivo je sto je od samog pocetka fiziologije , a narocito od sredine XIX veka , kada je prikupljeno dovoljno cinjenicnog materijala o dejstvu fizioloski aktivnih materija , fiziologe , farmakologe i lekare uopste interesovao problem mehanizma , tj. nacina dejstva bioloski aktivnih materija , a narocito bakterijskih i drugih toksina i lekova na celije.
Fosfatna jedinjenja bogata energijom . Mada je problem i pored veoma obimnih ispitivanja dobrim delom jos uvek otvoren , danas je u savremenoj fiziologiji i farmakologiji opstttthvacena teorija o celijskim hemijskim receptorima , tj. odredjenim hemijskim grupama u pojedinim jedinjenjima ( sastojcima ) celija za koje se fizioloski aktivne materije vezuju prilikom svoga dejstva na celiju.
Fosfatna jedinjenja bogata energijom .Teorija o hemijskim receptorima potice od tumacenja P. Ehrlicha o mehanizmu dejstva toksina (otrova) na celije , po kome svaka toksicna supstancija u svome molekulu sadrzi odredjenu hemijsku grupu – haptoforna grupa – kojom se vezuje za odredjene hemijske grupe u celijama .
Fosfatna jedinjenja bogata energijom . Tek posle vezivanja toksicna (toksforna) grupa ispoljava svoje dejstvo .
Fosfatna jedinjenja bogata energijom . Ovo svoje shvatanje Ehrlich je formulisao svojom poznatom izrekom Corpora non agunt nisi fixata .
Danas je , zahvaljujuci radovima velikog broja naucnika , a narocito H. Dalea , W.Feldberga , de Jonga i Ariensa ,formulisana koncepcija o celijskim hemijskim receptorima .
Fosfatna jedinjenja bogata energijom . Hemijski receptori celija su specificne grupe molekula , molekuli ili delovi molekula u celijama za koje se vezuju fizioloski aktivne supstancije preko kkspoljavaju svoje dejstvo .
Fosfatna jedinjenja bogata energijom . Ovi receptori se nalaze u raznim delovima ceija : celicnoj membrani , citoplazmi , mitohondrijama , jedru itd.
Fosfatna jedinjenja bogata energijom . Vrlo cesto su receptori delovi molekula pojedinih enzima , kao osto je slucaj sa acetil-holin-esterazom celicne membrane , ciji deo molekula sluzi kao mesto vezivanja acetil-holina , ili sa gvozdjem porfirinske grupe koenzima Varburgovog fermenta , za koju se vezuju otrovi celijskog disanja CO (za Fe++) odnosno HCN (za Fe+++) .
Posle vezivanja bioloski aktivne supstancije ispoljavaju svoje dejstvo : acetil-holin izaziva povecanu provodljivost i depolarizaciju celicne membrane , a CO ili HCN inhibiraju aktiviranje kiseonika .
Aktiviranje kiseonika u celijskom disanju . Ponekad , kao sto je slucaj sa acetil-holinom , ferment na kome se nalazi receptorna grupa razlaze bioloski aktivnu materiju , cime prestaje njeno dejstvo , dok se u nekim drugim slucajevima bioloski aktivne supstancije otklanjaju dejstvom drugih enzima , koji ih razlazu , najcesce oksidisu ,vezuju sa drugim jedinjenjima (glukuronskom ili sumpornom kiselinom) , ili se otklanjaju izlucivanjem iz organizma .
Aktiviranje kiseonika u celijskom disanju . Mehanizam vezivanja hemijskih receptora sa bioloski aktivnim supstyancijama je veoma razlicit :
Veze tipa soli , estarske veze , vodonicne veze itd. Razumljivo je da je i cvrstina veza razlicita i da zavisi od mnogo faktora , kao sto su koncetracija aktivne supstancije , tip veze , brzina izlucivanja , brzina razlaganja odnosno neutralizovanja itd.
Aktiviranje kiseonika u celijskom disanju . Isto tako je i vreme trajanja veze aktivne supstance – hemijskih rastvora – veoma razlicito .
Aktiviranje kiseonika u celijskom disanju .Ukoliko je receptor ferment koji razlaaaau aktivnu suspstancu , onda vreme iznosi milisekund ( na primer kod acetil-holin-esteraze )ili koji sekund.
Fosfatna jedinjenja bogata energijom . U slucaju da se radi o supstancama koje blokiraju aktivne grupe vezujuci se cvrsto ( na primer CO ili HCN kod Varburgovog fermenta odnosno jedinjenja koja se vezuju za SH-grupe ) spoj je mnogo trajniji , pa cak i potpuno trajan.
Fosfatna jedinjenja bogata energijom . Iz recenog se lako moze zakljuciti da su aktivne grupe hemijskih receptora cesto sasvim odredjene strukture pokazuju afinitet prema bioloskoj aktivnoj supstanciji .
otuda proistice da citavi nizovi hemijski srodnih jedinjenja ispoljavaju vrlo slicno biolosko dejstvo , posto se vezuju za iste receptore .
Fosfatna jedinjenja bogata energijom . Zavisno od ostalog dela molekula , specificnost dejstva moze biti ralicita . Dejstva slicno acetil-holinu ispoljava niz drugih estara holina , samo u razlicitom stepenu .
Ako se dejstvo acetil-holina uzme kao 1 ,N-butiril-holin ce imati dejstvo 0.3 (bice 3 puta slabiji ), kapronil-holin potpuno neaktivan.
Fosfatna jedinjenja bogata energijom . U drugim slucajevima promena strukture bioloski aktivne materije cini da ta materija zadrzava sposobnost vezivanja za receptor , ali se menja nacin dejstva:
Mesto stimulatornog (nadrazajnog ) javlja se inhibitorno (paralisuce) dejstvo i obrnuto .
Dok su aminooksidaze receptori za dejstvo fenil-etil-amina , anfetamin , koji je otporan prema dejstvu ovog enzima i sa kojim gradi stabilnije jedinjenje , prestavlja inhibitor ovog enzima .
Sinteticki nonapeptid koji u sebi sadrzi samo jedan molekul tirozina vise i leucin umesto asparaginske kiseline predstavlja inhibitor hormona neurohipofize vazopresina .
Fosfatna jedinjenja bogata energijom . podrobniji podaci o ostalim fatorima koji uticu na reakciju bioloskih aktivnih jedinjenja i hemijskih receptora kao i o kvantitativnim odnosima izmedju aktivnih susptanca i inhiobitora , mogu se naci u literaturi posvecenoj problemu hemijskih receptora odnosno antimetabolika .
U sledecem clanku iz oblasti fiziologije ocekuje te detaljna prica o krvi – njenoj strukturi, sastavu, funkcijama i ogromnom znacaju za organizam. Saznaces kako krv doprinosi odrzavanju zdravlja, transportu kiseonika, odbrani tela i zasto je neophodna za zivot i opstanak svake celije.