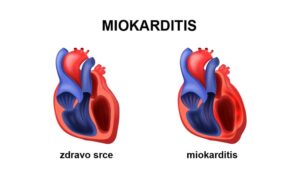
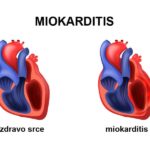
biohemijski sastav eritrocita
Biohemijski sastav eritrocita
Biohemijski sastav eritrocita: Sastoje se od 70% vode i hemoglobina (140–160 g/l), bez jedra, sa lipoproteinskom membranom i citoplazmom.
Eritrociti se sastoje od oko 70% vode, dok preostalu kolicinu cini uglavnom hemoglobin cija se koncentracija krece od 140g/l (kod zena) do 160g/l krvi (kod muskaraca) i koji je respiratorni pigment. Celija se sastoji od membrane (lipoproteinske strukture) i citoplazme bez jedra i vecine drugih celijskih organela
Bihemijski sastav eritrocita detaljnjje izlozen u biohemiji. Ovde cemo ga stoga samo rezimirati. Eritrociti pored vode (60-67) sadrze malu kolicinu malu kolicinu soli, pri cemu je nazniji katjon kalijum , a anjoni hloridi , fosfati i bikarbonati.
Metabolicke odlike srcanog misica 26
Metabolicke odlike srcanog misica Metabolicke odlike srcanog misica .Srcani misic (miokard) ima specificne metabolicke odlike...

Bioelektricne pojave u srcu 26
Bioelektricne pojave u srcu EKG elektrokardiogram Bioelektricne pojave u srcu omogucavaju stvaranje i sirenje elektricnih...

Fizioloske osobine srcanog misica 26
Fizioloske osobine srcanog misica Fizioloske osobine srcanog misica (miokarda)omogucavaju neprekidan rad srca kroz automatsko stvaranje...
Od organskih sastojaka u crvenim krvnim zrncima nalazimo trag ugljenih- hidrata , lipoide (slobodni i esterifikovani holesterin) , fosfatide (kefaline, lecitine, sfingomilejine) i belancevine.
Lipoidi se uglavnom nalaze vezani s proteinima u obliku lipoproteida . Medju belancevinama gde pored lipoproteida spadaju nukleoproteidi i nerastopljivi proteini strome , posebni znacaj ima hemoglobin , kako po kolicini tako i po fizioloskom znacaju.
Hemoglobin cini oko 32% ukupne tezine eritrocita , odnosno preko 90% njihove suve tezine.
Hemoglobin – osobine i fizioloska uloga
Iz biohemije poznato da je hemoglobin slozena belancevina , hromoproteid , koja se sastoji od proste belancevine globina , molekulske tezine oko 68 000 , i prosteticke grupe , porfirinskog derivata hema. Za svaki molekul globina vezano je po 4 molekula hema.
Hem je porfirinski derivat – feroprotoporfirin IX , u kome je dvovalentno fero-gvozdje vezano sa dve prave i dve kovalentne veze za azotne atome protoporfirina .
Sa preostale dve kovalence gvozdje je vezano za globin. U prisustvu kiseonika , kada su ostvareni i drugi uslovi o kojima ce kasnije biti reci , oslobadja se jedna kovalenca koja je bila gvozdje hemoglobina.
Vec na ovom mestu isticemo da gvozdje u oksi hemoglobinu , posle vezivanja kiseonika kovalentnom vezom ostaje i dalje u fero-obliku , znaci dvovalentno.
Posto svaki molekul hemoglobina sadrzi u sebi 4 hema , pa prema tome i 4 atoma gvozdja , to ce vezati ukupno 4 molekula kiseonika.
O uslovima vezivanja kiseonika za hemoglobin i postanku oksihemoglobina , kao i o disocijaciji oksihemoglobina , tj. otpustanju kiseonika i prelasku u hemoglobin , bice vise reci u fiziologiji disanja.
Ovde samo napominjemo da se oksihemoglobin stvara pri dovoljno visokom gasnom pritisku kiseonika, odnosno u plucima gde se kiseonik nalazi u smesi sa drugim gasovima vazduha , pri dovoljno visokom parcijalnom pritisku O2.
Ovako vezani kiseonik prenosi krv iz plucnih kapilara u tkiva , gde je parcijalni pritisak kiseonika vrlo nizak , usled cega se oksihemoglobin disosuje i oslobadja kiseonik , koji iz krvi tkivnih kapilara prelazi u celije i tamo ucestvuje u sidacionim procesima.
Tipovi hemoglobina
Uporednim proucavanjem fizicko-hemijskih i biohemijskih osobina hemoglobina raznih kicmenjaka konstatovano je da svi do sada poznati hemoglobini , bez obzira na poreklo , imaju identican hem , dok se razlikuju po osobinama globina.
Mada se kiseonik vezuje za hem , ipak od osobine globina proces vezivanja i otpustanja kiseonika. Danas se zna da razlike izmedju globina pojedinih hemoglobina raznih kicmenjaka zavise od procenta pojedinih aminokiselina , kao i od prostorne konfiguracije globinovog molekula.
Novija ispitivanja su pokazala da i hemoglobin kod iste vrste , na primer kod coveka , nije uvek isti . Tako je kod coveka i velikog broja sisara konstatovano je da kod fetusa , a delom i novorodjencadi postoji fetalni , koji se po nizu odlika ralikuje od odraslih osoba.
Osim toga kod bolesnika sa razmin vrstama anemija ( malokrvnosti) , konstatovano je prisustvo posebnih „patoloskih hemoglobina“.
Sustina da izmedju ovih takodje pociva na razlicitom sastavu peptidnih lanaca globina i njihovoj konfiguraciji , a razlike se manifestuju u elektroforetskoj pokretljivosti , otpornosti prema alkalijama , ponasanju pri hromatografiji i fizioloskim odlikama: lakoci vezivanja i otpustanja kiseonika.

Sadrzaj hemoglobina
Tehnika odredjivanja sadrzaja , u krvi opisana je u fizioloskim i biohemijskim praktikumima . Zdrave osobe sadrze prosecno 14-17 g u 100g krvi ( muskarci od 15-17 , a zene 14-16 g%), odnosno 79-80% po Sahliju. Unovije vreme sadrzaj se uglavnom izrazava g%.
Derivati hemoglobina
Relativno labilno jedinjenje i citav niz hemijskih supstancija moze s njim reagovati i pretvoriti ga u derivate koji vise nemaju sposobnost vezivanja i prenosenja kiseonika.
Kako se takvi derivati mogu stvarati i u organizmu pri razlicitim trovanjima , to cemo pomenuti najvaznije od njih. Jedan od najprostijih i najsigurnijih nacina razlikovanja pojedinih derivata je spektroskopsko razlikovanje , posto hemoglobin i njegovi derivati apsorbuju svetlosne znake odredjene talasne duzine , sto se manifestuje pojavama tamnih apsorpcionih pruga u odredjenom delu spektra.
Opis spekroskopa i tehnika pripremanja pojedinih derivata izlozeni su u biohemijskim i fizioloskim praktikumima. Oksihemoglobin , ciji se rastvor dobija hemolizom male kolicine krvi destilisanom vodom , pokazuje da dve apsorpcione grupe u zuto-zelenom delu spektra ( u odlasti D i F Frauhoferove linije).
Ukoliko se rastvoru doda nekoliko kapi Stockesovog reagensa ( amonijacni rastvor ferotartarata), dolazi do oduzimanja kiseonika od oksihemoglobina , cime se dobija hemoglobin , u obicnom zivotu hemijski nepravilno nazvan „redukovani hemoglobin“( posto je gvozdje i u njemu , kao i u oksihemoglobinu , dvovalentno).
Karboksihemoglobin – gvozdje pokazuje vrlo veliki afinitet prema ugljen monoksidu – 300 puta ego prema kiseoniku.
Otuda i pri vrlo malim koncetracijama CO u vazduhu (0.01%) za vrlo kratko vreme najveci deo eritrocita biva zasicen ugljen-monoksidom , pri cemu postaje derivat , u kome je CO takodje vezan kovalentno , samo je ta veza mnogo cvrsca .
Ovim krv gubi sposobnost prenosenja kiseonika do tkiva i stoga u tkivima dolazi do nedostatka kiseonika i asfiksije ( gusenja ). Mada je karoksimohemoglobin mnogo stabilniji od oksihemoglobina , ipak u prisustvu kiseonika i do njegove disocijacije i otpustanja CO .
Stoga pri lecenju trovanja ugljen-monoksidom otrovanima daje se gasna smesa bogatija kiseonikom od obicnog vazduha , kako bi se olaksalo otpustanje CO i tkivima stavlja na raspolaganje sto veca kolicina kiseonika .
Spektar odlikuje se prisustvom dve apsorpcioe pruge u zuto-zelenom delu spektra , koje su pomerene malo udesno – prema ljubicastoj boji , u poredjenju sa spektrom oksihemoglobina .
Stokesov reagens ne redukuje karboksihemoglobin , te posle njegovog dodavanjne dollldo pojave jedne apsorpcione pruge , na mesto dve .
Methemoglobin je oksidacijom gvozdja hema, pri cemu ovo gvozdje prelazi u trovalentni feri-oblik. Za slobodnu valencu se obicno vezuje OH- grupa .
Kupite knjigu

Ostali derivati . Pri trovanju sumpor-vodonikom u organizmu moze postati hemoglobinov derivat sulfohemoglobin , a pri trovanju azotovim oksidima nitrozehemoglobin. I ovi derivati imaju karakteristicne apsorpcione pruge na spektru.
Uloge hemoglobina – ima niz vaznih fizioloskih uloga. Kanajvazniju , nesumljivo treba pomenuti prenosenje kiseonika iz pluca do tkiva , cime se celije snabdevaju potrebnom kolicinom kiseonika. Druga uloga je u prenosenju ugljen-dioksida.
Za slobodne amino-grupe globina vezuje se ugljen-dioksida , pri cemu postaje karbaminohemoglobin. Treba istaci da pri vezivanju kiseonika za gvozdje hema i postanku oksihemoglobina dolazi do disocijacije karbaminohemoglobina i otpustanja CO2.
Oko 40% ukupnog ugljen-dioksida venske krvi vezano je u obliku karbaminohemoglobina . Sledeca vazna uloga je njegova puferska uloga i ucesce u odrzavanju acido-bazne ravnoteze krvi.
Fizicko-hemijska ispitivanja su pokazala da sui hemoglobin i i-hemoglobin organske kiseline. Pri tom je hemoglobin vrlo slaba kiselina , slabija od ugljene kiseline , dok je oksikohemoglobin znatno jaca kiselina – jaca od H2CO3 .
Na ovoj razlici i pociva puferska uloga . U plucima vezivanjem O2 od hemoglobina postaje oksihemoglobin , kiselina jaca od ugljene . Okkmoglobin reaguje sa kalijum-bikarbonatom eritrocita i iz njega istiskuje ugljenu kiselinu , koja je pod uticajem fermenata karbonhidraze razlaze na CO2 i H2O, a CO2 izbacuje iz plucne mehurice i odavde , procesom izdisanja , u atmosferu.
Oksihemoglobin za sebe vezuje kalijum oduzet od KHCO3 , pri cemu postaje kalijum-oksihemoglobinat .
U tkivnim kapilarima kalijum-oksihemoglobinat najpre otpusta kiseonik koji prelazi u tkiva i trosi se u procesima celicnog disanja . Istovremeno iz tkiva u kapilare prelazi ugljen-dioksid koji je postao u toku prometa materijeee, ulazeci kako u krvnu plazmu tako i u eritrocite .
Tu se CO2 rastvara u vodi i daje ugljenu kiselinu . Ugljena kiselina koja je na ovaj nacin postala u eritrocitima jaca je od hemoglobina , te od njega oduzima kalijum , pri cemu postaje vrlo slaba kiselina H-Hb i kalijum karbonat.
Ovom svojom funkcijom obezbedjuje odrzavanje stalnosti elektrohemijske reakcije krvi i pored toga sto se u plucnim alveolama gubi znatna kolicina ugljen-dioksida i oslobadja znatna kolicina alkalija iz bikarbonata , a u tkivnim kapilarima iz celija u krv dolazi znatna kolicina ugljen-dioksida i postaje ugljena kiselina.
Medjutim , zahvaljuci vezivanju alkalija (kalijuma) za oksihemoglobin u krvi plucnih kapilara i njihovom oslobadjanju iz kalijum-hemoglobinata u tkivnim kapilarima , elektrohemijska reakcija krvi se prakticno ne menja : snizava se svega 0.02 pH u arterijskoj krvi.
Stvaranje eritrocita
Eritrociti se stvaraju u crvenoj kostanoj srzi pljosnatih kostiju (sternum , rebra i lobanjske kosti), u sujerastim kostima i u epifizama dugih kostiju ; kod dece i mladjih osoba erotrociti se stvaraju i u crvenoj kostanoj srzi dugih kostiju , koja se , kao sto je poznato , kasnije pretvara u zutu srz, dok se u fetalno doba eritrociti stvaraju i u slezeni i jetri.
Pri vecem gubitku krvi i nekim oboljenjima krvnog sistema moze i u postfetalnom dobu kod dece, pa cak i odraslih , doci do stvaranja eritrocita u jetri i slezeni.
Tok stvaranja eritrocita od osnovne celije hematopoeticne loze hemohistioblasta – izlozen je u udzbenicima histologije.
Fatkori stvaranja eritrocita
Za pravilno stvaranje eritrocita pored neostecene funkcionalne sposobnosti kostane srzi neophodno je unosenje dovoljne kolicine gvozdja , teskih metala bakra i mangana , koji omogucuju redukciju feri-gvozdja u fero-oblik i njegovo ugradjivanje u molekul hema , esencijalnih amino-kiselina , kao i skoro svih vitamina .
Od narocito neposrdne vaznosti za stvaranje i sazrevanje eritrocita i sintezu hemoglobina su vitamini B12 , C i folna kiselina.
Regulacioni faktori. Mnogobrojna ispitivanja su pokazala da je nedostatak kiseonika u organizmu (anoksemija) jedan od najmocnijih najvaznijih faktora koji stimulisu aktivnost kostane srzi i stvaranje eritrocita .
Pomenucemo kao primer dva slucaja koji dovode do anoksemije . Prvi je pri boravku na visokim planinama iznad 3000m , gde je usled snizenog atmosferskog pritiska smanjeno snabdevanje kiseonikom.
Organizam se na ove uslove adaptira pojacanim stvaranjem eritrocita , tako da se broj u perifernoj krvi moze vecati za 2 i vise miliona iznad prosecnog u 1mm3 .
Drugi slucaj je gubitak krvi pri iskrvarenjima . Eksperimentalno je konstantovano da ubrizgavanje seruma zivotinja stimulise pojacano stvaranje eritrocita i pojavu mladih oblika (retikulocita) u perifernoj krvi normalnih zivotinja.
Poseeem preparativnom metodom je iz seruma iskrvarenih zivotinja , kao i iz mokrace i niza organa (bubrezi i dr.) , izolovana supstancija eritropoetin , koja stimulise aktivnost kostane srzi.
Mesto stvaranja i biohemijska priroda eritropoetina nisu jos rastavljeni , ali je sporno da se on stvara kao reakcija na opstu ili lokalnu anoksemiju i da stimulise stvaranje eritrocita u kostanoj srzi.
Zivot i raspadanje eritrocita
Eritrociti prosecno zive 3 do 4 meseca ( oko 90 d0 140 dana) , polse cega dolazi do njihovog raspadanja u slezini i retikuolendotelnim elementima (Kupferovim celijama) jetre.
Najcesce retikuloendotelni elementi uvlace u sebe (fugocituju)ostecene i ostarele eritrocite , posle cega pod uticajem fermenta tih celija dolazi do dezintegracije (razaranja ) eritrocita i razlaganja njihovih sastojaka .
Pri tom od hemoglobina postaju zucne boje, o cijem ce postanku biti reci pri proucavanju sastava zuci.
Pored opisanog fizioloskog raspadanja , pod uticajem raznih hemijskih otrova (fenil-hidrazin , benzol i niz drugih )i bioloskih faktora (zmijski otrovi , otrovi insekata , bakterijski toksini , aglutinini idr.)moze moze doci i do hemolize u opstem krvotoku.
Jos je ne razjasnjen mehanizam hemolitickog dejstva slezine kod splenomegalija , oboljenja pracenog povecanjem slezine , kada se javlja pojacano raspadanje eritrocita , koje moze dovesti do teske anemije.
Neki autori smatraju da obolela slezina izlucuje hemoliticke supstancije koje u opstem krvotoku dovode ostecenja eritrocccc hemolizom ,,,,
U prilog ovom shvatanju govorila bi smanjena rezistencija eritrocta kod splenomegalije . Medjutim , hemoliticke supstancije slezine nisu dobivane u koncetrovanom i preciscenom stanju .
Postoji i glediste da i normalna slezina sadrzi izvesnu kolicinu hemolitickih supstancija, posto davanje ekstrakata slezine po nalazu nekih autora dejstvuje hemoliticki. S druge strane , kstirpacija (vadjenje) slezine dovodi , bar prolazno ,do povecanja broja eritrocita.
Leukociti bela krvna zrnca
Druga grupa uoblicenih elemenata krvi su leukociti ( bela krvna zrnca ). Svezi jos zivi leukociti se mogu uzeti na vise nacina. Prvi je centrifugovanje citratne krvi i uzimanje povrsnog belicastog castog sloja , koji u obliku tanke skrame pokriva eritrocite , a drugi je izdvajanje eritrocita centrifugovanjem citratne krvi malom brzinom (800 – 1000 obrtaja u minutu) i adsorpcijom leukocita iz plazme na plasticnu smolu amberlit, sa koje se leukociti mogu elulirati pogodnim puferom.
Na nativnom preparatu leukociti izgledaju kao male prozracne celije nepravilnog oblika , koje pokazuju ameboidno kretanje ispustajuci pseudopodije (nastavke citoplazme).
Leukociti se obicno posmatraju na trajnom , fiksiranom i obojenom preparatu. Bojenje moze biti razlicito , ali se u fizioloskim i klinickim labaratorijama najcesce primenjuje bojenje azur-eozinom po Gimzi ( Giemsa -Romanowski), metilinskim plavimi eozinom u metil-alkoholu po May- Grunwaldu ), ili kombinovano po Papheimu .
Tehnika bojenja opisana je u odgovarajucim praktikumima. Na fiksiranom preparatu leukociti imaju okugao oblik , precnik im je , zavisno od vrste , od 8 (limfociti ) do 25 mikrona (monociti) , sa jasno izrazenim jedrom razlicitog oblika i citoplazmom, koja je opet razlicita , zavisno od vrste leukocita – amforna ili granulirana .
Broj leukocita
Brojanje leukocita vrsi se slicno brojanju eritrocita. U manji melanzer uvuce se krv do oznake 0.5 ili 1.0 a zatim Turcov rastvor* do oznake 11 , pa se obtanjem dobro izmesa .
Prisutna sircetna kiselina razara eritrocite , a boja oboji jedra leukocita i trocita , s tim sto se brojanje vrsi u celoj komori ( svih 400 kvadratica).
Prosecan broj leukocita kod coveka krece se izmedju 6 -8000 u 1mm3.
Promene broja leukocita . Broj leukocita pokazuje prilicno velike varijacije vec pod fizioloskim okolonostima. Tako kod zdravih osoba dolazi do povecanja broja leukocita posle uzimanja hrane , za vreme varenja , u toku fizickih naprezanja , a kod zena za vreme menstrualnog ciklusa i bremenitosti.
Kod raznih oboljenja dolazi do jos vecih varijacija broja leukocita , i to bilo do povecanja broja leukocita – leukocitoze , ili do smanjenja leukopenije .
Detaljni podaci o kretanju broja leukocita kod raznih oboljenja izlazu se u udzbenicima pataloske fiziologije i unutrasnjih bolesti. Ovde samo napominjemo da se broj leukocita povecava kod svih gnojnih procesa i mnogih oblika maligne proliferacije leukopoeticnih tkiva – kod leukoza.
Broj leukocita je smanjen kod trbusnog tifusa , a narocito posle ostecenja kostane srzi raznim industrijskim otrovima i jonizujucim zracenjima ( rendgenski zraci , radioaktivna zracenja itd.).
Vrste i podela leukocita
Prema poreklu , obliku jedra i izgledu citoplazme leukociti se dele u dve osnovne grupe : na leukocite mijeloidne loze ili granulocite , koji se stvaraju u kostanoj srzi i na leukocite limfoidne loze , koji se stvaraju u limfnom tkivu : limfni cvorovi i slezina .
Leukociti mijeloidne loze – granulocidi – stvaraju se u kostanoj srzi. Odlikuju se raznjevitim jedrom , ciji broj raznjeva zavisi od stvarnosti leukocita : sasvim mladi leukociti mijeloidne loze imaju neraznjevito „stapicasto“ jedro , a zatim broj raznjeva raste sa staroscu leukocita.
Citoplazma leukocita mijaloidne loze je granulisana , pri cemu se granulacije boje bilo neutralnim ili kiselim odnosno baznim bojama.
Prema afinitetu granulacija za boje leukociti mijeloidne loze dele se u tri grupe: neutrofilni , cije se granulacije boje neutralnim bojama , eozifilni ili acidofilni , cije su granulacije boje kiselim bojama (eozinom) , i bazofilni , cije se granulacije boje baznim bojama.
Leukociti limfoidne loze -agranulociti – stvaraju se u limfoidnom tkivu – limfnim cvorovima i slezini Citoplazma im je bez granulacija .
Dele se u dve grupe : a) limfociti , male celije precnika 6-8 mikrona , sa malo izrazenom citoplazmom u obliku uskog pojasa , oko jedra ; b)monociti krupne celije precnika oko 20 mikrona.
Leukocitna formula. Leukocitna formula pokazuje brojni odnos izmedju pojedinih vrsta leukocita . Odredjuje se na taj nacin sto se na obojenom preparatu („razmazu krvi“) odredi brojni odnos pojedinih vrsta leukocita na dovoljnom broju (200-300) leukocita.
Leukocitna formula kod coveka glkasi : neutrofilnih leukocita 60-70% , eoizinofilnih 2-4% , bazofilnih 0.5% , limfocita 25-30% , i monocita 6-8%.
Leukocitna formula kod pojedinih oboljenja poazuje takodje karakteristicne promene . Ne ulazeci u detalje koji se mogu naci u patoloskoj fiziologiji , isticemo da se pri gnojnim procesima i u akutnoj fazi mnogih infektivnih oboljenja javlja neutrofilija – povecanje broja neutrofilnih leukocita.
Broj eozinofilnih leukocita je povecan kod mnogih alergicnih stanja i kod invazije crvenim i drugim parazitima.